
JSER Policies
JSER Online
JSER Data
Frequency: quarterly
ISSN: 1409-6099 (Print)
ISSN: 1857-663X (Online)
Authors Info







- Read: 13496
Анастас ЛАКОВСКИ
ЗНАЧЕЊЕТО НА ГЕНОМСКИОТ ЗАПИС (GENOMIC IMPRINTING) ВО ХУМАНАТА ГЕНЕТИКА И ДЕФЕКТОЛОГИЈАТА
Увод
Во рамките на хуманата популација често се јавуваат својства (заболувања) со генетска основа чија што трансмисија не подлежи на Менделовите закони. Типичен и најпознат пример на отстапување од Менделовите закони на трансмисија секако се полово врзаните својства, односно X-врзано рецесивните својства, покрај нив отстапување од Менделовите закони пројавуваат, новонастанатите спонтани мутации, реорганизациите на хромозомите, зголемениот број на триплетни повторувања, митохондријално условените својства и секако својствата што се јавуваат заради појавата на геномскиот запис (genomic imprinting).
Отстапувањето на полово врзаните својства од Менделовите закони на трансмисија е општо познато. Корелирано пак со одново откриеното значење на триплетните повторувања, на пример фрагилизациите на X-хромозомот како една од етиолошките основи на пројавноста на менталната ретардација нашите позавања за трансмисијата на полово врзаните својства ги редефинираме и велиме дека фрагилниот X-синдром е X-врзано својство со атипична трансмисија.
Повеќето генетски условени својства (заболувања) претспоставуваат ново настанати мутации што не се пренесуваат од генерација на генерација па оттаму и не подлежат на Менделовите закони на трансмисија.
Реорганизациите на хромозомите, како од нумерички така и од структурен аспект како што е познато не мора да се наследуваат, на пример трисомијата на 21-от хромозом што може да се јави како последица на нераздвојувањето и што ќе резултира со една од формите на Dowen-овиот синдром.
Триплетните повторувања исто така, како што веќе подвлековме се етиолошка основа за многу заболувања што во трансмисијата манифестираат карактеристично отстапување од Менделовите закони.
Изучувањето на појавата на триплетните повторувања е во подем и благодарение на моќта на современата молекуларна генетика нивното значење с¢ повеќе се потенцира и с¢ повеќе се осознава нивната улога во рамките на човековиот геном. Покрај познатиот фрагилен X-синдром познато е дека во генетската етиологија на Хантингтоновата кореа (Huntington's chorea) исто така, иако не секогаш!, лежи триплетно повторување.
Митохондријалните заболувања во својата есенција манифестираат карактеристично отстапување од Менделеевите закони на наследување и на некој начин симулираат X-врзана трансмисија пред с¢ заради бактериолошкото потекло на митохондријалната ДНК и нејзината застапеност само во јајцевите клетки.
Геномскиот запис е појава на која што £ се посветува внимание заради дивергенцијата во експресијата на својствата што ги условуваат геномски импринтираните гени и заради се поголемиот број на својства што се откриени благодарение на молекулараната генетика, а за кои се верува дека во нивната пројавност значително партиципира геномскиот запис.
Што е геномски запис (genomic imprinting)
Современата литература од областа на хуманата генетика појавата на геномскиот запис ја дефинира како варијација во експресијата на генот во зависност од тоа дали мутираниот (заболениот) ген е наследен од таткото или од мајката.
Појавата не геномскиот запис најдобро се објаснува на примерот на различната експресија на структурните реорганизации на хромозомите, најчесто делециите, во зависност од тоа дали делетираниот хромозом води потекло од мајката или од таткото. Секако дека кај делециите не зборуваме за ген туку за гени што се лоцирани во рамките на регионот на хромозомот што е делетиран.
Во хуманата популација познати с¢ повеќе синдроми што етиолошки се поврзуваат со делеции на определени региони на хромозомот и манифестираат определено ниво на животна конкурентност, најчестите синдроми асоцирани со делеции можат да се видат од следната табела:
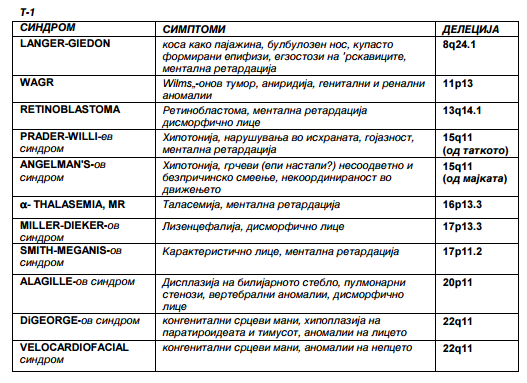
Карактеристични синдроми како последица
на делеции на различни хромозоми
Хромозомска основа на геномскиот запис
Може да се претпостави дека гените што се поставени во рамките на делетираниот регион имаат 50% веројатност да доживеат фенотипсска експресија, но понекогаш тоа не е така. Некои гени, во зависност од тоа дали водат порекло од мајчиниот или од татковиот алел не доживуваат еднаква фенотипска експресија. Во вакви случаи најчесто само алелот од мајчиниот хромозом или само алелот од татковиот хромозом ќе доживее експресија, барем во текот на раниот развој. Доколку делетираниот алел е тој што е нормално активен, генот ќе биде функционално отсутен, и покрај тоа што една копија е присутна. Од табелата, Т-1, се забележува дека генетската основа на различни синдроми PRADER-WILLI-ев синдром и ANGELMAN'S-ов синдром, DiGEORGE-ов синдром и VELOCARDIOFACIAL синдром, е во делеција на идентичен хромозом но со различно потекло.
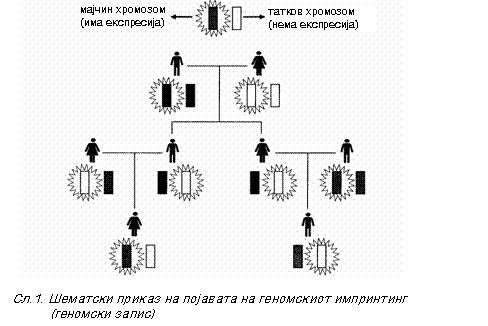
Појавата на геномскиот запис (genomic imprinting) најнапред е изучувана кај цицачите во опитите со глувците (стаорците)4, при што е воочено дека ембрионите добиени со дупликација или на мајчиниот или на татковиот хромозомски сет биле животно неспособни (Сл. 2).
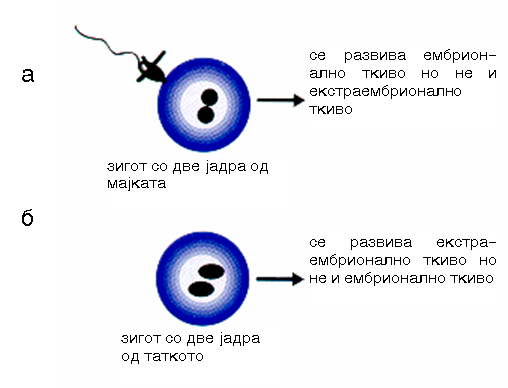
Сл. 2 Микрохируршка јадрена трансформација на зигот на глушец:
а) со две мајчински јадра;
б) со две таткови јадра
Мора да се подвлече дека животната инкомпатибилност не се јавува поради хомозиготноста, затоа што, глувците, ионаку настануваат од линии што нормално се хомозиготни! Оттаму сосема е јасно дека за продуцирањето на животот на овој начин е неопходен хромозомскиот сет од секој родител.
Со помош на сетови на хромозоми со балансирани транслокации, добиени се глувци што двете копии на еден хромозом или пак негов регион ги наследиле од еден родител. Овие експерименти покажале дека некои региони на хромозомите се геномски импринтирани додека други не.
Сосема друг систем за изучување на појавата на genomic imprinting е т.н. трансгенетски глушец. Имено, надворешни фрагменти на ДНК можаат да интегрираат со импринтираните региони на хромозомите на глушецот. Трансмисијата на овие трансгени од едниот родител ќе резултира со експресија на генот додека трансмисијата на трансгенот од другиот родител нема да доживее фенотипска експресија. Imprinting-от се јавува во герминативната линија на секоја генерација.
Откриено е дека импринтираните гени трпат метилација на некои CpG места, што упатува на заклучок дека метилацијата можеби е инволвирана во супресијата на активноста на импринтираните региони.
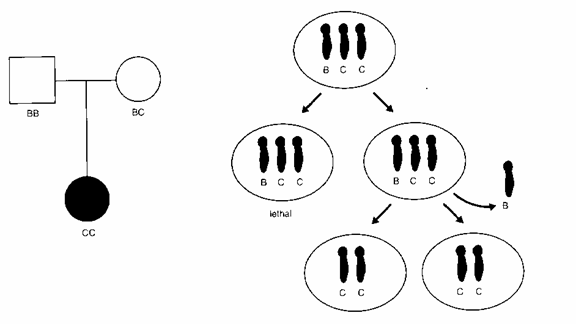
Ефектите на појавата на genomic imprinting кај човекот најнапред се инволвирани во разјаснувањето на проблемите кај единките заболени од цистична фиброза, кои меѓу другото манифестираат тешка развојна и ментална ретардација. Кај овие единки е докажано дека мутираниот ген за цистична фиброза го наследиле меѓу другите гени поставени вдолж 7-от хромозом само од едниот родител што може да се види од следната слика:
Сл. 3. Мајчинска изодисомија што резултира
со цистична фиброза кај детето
Означени се хаплотиповите за полиморфниот маркер што тесно се поврзува со цистичната фиброза. Заболеното дете кое покрај кусиот раст манифестира цистична фиброза двете копии од маркерот ги наследило од мајката што е носител на цистична фиброза. Татковиот алел Б не е застапен кај детето.
Оваа појава во хуманата генетика е наречена унипарентерална дисомија. Се чини дека унипарентералната дисомија е последица на елиминацијата на еден од хромозомите при трисомичните концепции. Трисомијата 7 е аконкурентна со животот, но доколку рано во ембрионалниот развој еден од трите 7 хромозоми ќе се изгуби поради nondisjuntion тогаш се реставрира нормалниот хромозомски сет. Доколку парот на 7-от хромозом што останал е добиен од ист родител (татко или мајка), настанува унипарентерална дисомија што секако ќе има фенотипски последици доколку хромозомите содржат импринтирани региони.
Ефектите на појавата на genomic imprinting може да се воочи кај делетивните синдроми. На пример Prader-Willi-евиот синдром и Angelman- овиот синдромот се дистинктивни заболувања што се асоцирани со делеција на ист регион на 15-от хромозом. Доколку делецијата е на татковиот 15-ти хромозом фенотипски ќе се изрази како Prader-Willi-ев синдром доколку пак делецијата е на мајчиниот 15-ти хромозом ќе се изрази како Angelman-ов синдром.
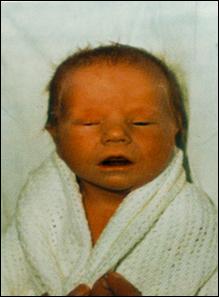
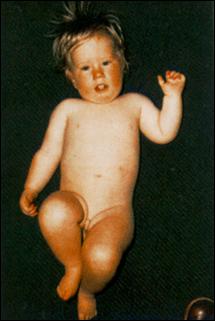
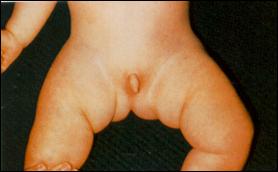
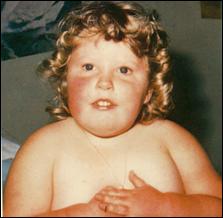
Кај некои случаи на Prader-Willi-евиот и Angelman-овиот синдромот цитогенетски не се докажува делеција но сепак бидејќи постои унипарентерална дисомија на татковата копија на 15-от хромозом се јавува Angelman-овиот синдромот ако пак постои унипарентерална дисомија на мајчината копија на 15-от хромозом се јавува Prader-Willi-евиот синдром. Веројатно, во односниот регион се јавуваат импринтирани гени при што некои се изразуваат ако се поставени на татковиот а некои ако се поставени на мајчиниот 15-ти хромозом.
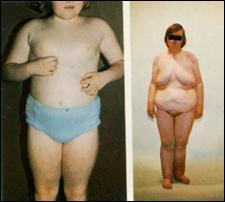
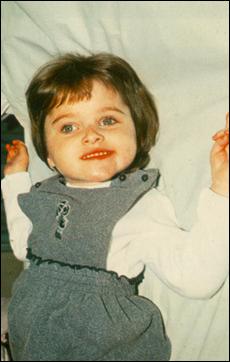
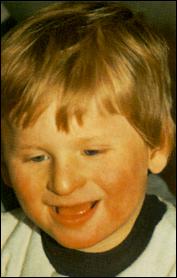
Сл. 4. Карактеристични симптоми на Prader-Willi-eviot синдром7
 |
 |
![]()
 |
 |
![]()
Вистинското ниво на фенотипската експресија на појавата на genomic imprinting-от сe уште не е спознаено. Постои теоретски пристап што укажува дека ограничениот плацентарен мозаицизам може да настане поради нераздвојување на трисомичните концепции што може да резултира со соодветни фенотипски последици доколку се јави унипарентерална дисомија. Исто така, постојат податоци дека потомството на фенотипски нормалните носители на балансираната транслокација е фенотипски абнормално без оглед што ја наследило истата избалансирана транслокација. Кај некои е утврдено дека обата хромозоми ги наследиле од еден родител, и оној со избалансираната транслокација и нормалниот, што резултира со унипарентерална дисомија. Кај други пак е утврдено дека имаат делеција што не е застапена кај родителот, што упатува на нестабилност на реорганизираниот хромозом. Сето ова упатува на заклучок дека вистинското ниво на фенотипските последици на хромозомските абнормалности воопшто, не е целосно утврдено.
Согласно со развојот на сознанијата од областа на хуманата генетика, дефектолозите како носечки партиципиенти во процесот на генетичкото советување треба континуирано да ги следат и осознаваат новините во областа на хуманата генетика. Само на тој начин, непосредно може да се придонесе во превенирањето на репојавноста на повеќето генетски но и наследно условени состојби следени со определени видови на попреченост во развојот. Превенирањето во многу би придонесло не само од аспект на намалувањето на нивото на општествените потреби за материјални издатоци неопходни за социјализација на овие лица, туку, корелирано со резултатите во областа на молекуларната генетика би можело да ги насочи активностите во примената на т.н. генотерапија.
ЛИТЕРАТУРА:
- Plomin R, DeFries C. J., McCl;earn E. G., Rutter M., BEHAVIORAL GENETICS, Third Edition, W.H. Freeman and Company, New York (1997) 22-26
- Warren S T., Nelson D l., ADVANCES IN MOLECULAR ANALYSIS OF FRAGILE X SYNDROME, JAMA, No.271 (1994) 536-542
- Bruce R. Korf, HUMAN GENETICS, A problema-based aproach, Blackwell Science, Cabridge, Massachusetts, USA (1996) 196-203
- Surani, M.A., GENOMIC IMPRINTING: CONTROL OF GENE EXPRESION BY EPIGENIC INHERITANCE, Curr.Pin.Cell Biol .No.6 (1994) 390-395
- Klug,S.W., Xummings R.M., ESENTIAL OF GENETICS, Second edition, Prentice Hall, New Jersey (1996) 152-143
- Spance, J.E., Perciaccante R.G., Greig G.M., UNIPARENTAL DISOMI AS A MECHANISM FOR HUMAN GENETIC DISEASE, Am. Journal of Hum Genet. No. 42 (1988) 217-226
- 8, 9., Baraister, M., Winter,M., Acolour atlas of CLINICAL GENETICS, Wolf Medical Publications Ltd. London (1990) 62,53
- Vogel, F., Motulsky G.A., HUMAN GENETICS, Problem And Aproaches, Third, Completely Revised Edition, Springer-Verlag, Berlin, Heidelberg, New York (1997) 285-288
Follow Us
Share Us
Journal metrics
-
 SNIP 0.059
SNIP 0.059 -
IPP 0.07
-
 SJR 0.13
SJR 0.13 -
 h5-index 7
h5-index 7 -
 Google-based impact factor: 0.68
Google-based impact factor: 0.68
Indexed in





![]()
10 Most Read Articles
- PARENTAL ACCEPTANCE / REJECTION AND EMOTIONAL INTELLIGENCE AMONG ADOLESCENTS WITH AND WITHOUT DELINQUENT BEHAVIOR
- RELATIONSHIP BETWEEN LIFE BUILDING SKILLS AND SOCIAL ADJUSTMENT OF STUDENTS WITH HEARING IMPAIRMENT: IMPLICATIONS FOR COUNSELING
- EXPERIENCES FROM THE EDUCATIONAL SYSTEM – NARRATIVES OF PARENTS WITH CHILDREN WITH DISABILITIES IN CROATIA
- INOVATIONS IN THERAPY OF AUTISM
- DIAGNOSTIC AND TREATMENT OPTIONS IN AUTISTIC SPECTRUM DISORDERS – AN OVERVIEW
- AUTISM AND TUBEROUS SCLEROSIS
- THE DURATION AND PHASES OF QUALITATIVE RESEARCH
- REHABILITATION OF PERSONS WITH CEREBRAL PALSY
- REVIEW OF SPECIAL EDUCATION PROGRAMS IN JORDAN: CURRENT PRACTICES, CHALLENGES, AND PROSPECTS
- DISORDERED ATTENTION AS NEUROPSYCHOLOGICAL COGNITIVE DISFUNCTION